Какие вещества клетки называют органическими. Неорганические вещества клетки
Вода – один из самых основных компонентов живой клетки, составляет в среднем 70-80% массы клетки. В клетке вода находится в свободной (95%) и связанной (5%) формах. Помимо того, что она входит в их состав, для многих организмов это еще и среда обитания.
Роль воды в клетке определяется ее уникальными химическими и физическими свойствами, связанными главным образом с малыми размерами молекул, с полярностью ее молекул и с их способностью образовывать друг с другом водородные связи. Вода как компонент биологических систем выполняет следующие важнейшие функции:
1. Вода - универсальный растворитель для полярных веществ, например солей, сахаров, спиртов, кислот и др. Вещества, хорошо растворимые в воде, называются гидрофильными.
2. Молекулы воды участвуют во многих химических реакциях, например при гидролизе полимеров.
3. В процессе фотосинтеза вода является донором электронов, источником ионов водорода и свободного кислорода.
4. Неполярные вещества вода не растворяет и не смешивается с ними, поскольку не может образовывать с ними водородные связи. Нерастворимые в воде вещества называются гидрофобными.
5. Вода обладает высокой удельной теплоемкостью. Для разрыва водородных связей, удерживающих молекулы воды, требуется поглотить большое количество энергии. Это свойство обеспечивает поддержание теплового баланса организма при значительных перепадах температуры в окружающей среде.
6. Вода отличается высокой теплопроводностью, что позволяет организму поддерживать одинаковую температуру во всем его объеме.
7. Вода характеризуется высокой теплотой парообразования, т. е. способностью молекул уносить с собой значительное количество тепла при одновременном охлаждении организма. Благодаря этому свойству воды, проявляющемуся при потоотделении у млекопитающих, тепловой одышке у крокодилов и других животных, транспирации у растений, предотвращается их перегрев.
8. Для воды характерно исключительно высокое поверхностное натяжение. Это свойство имеет большое значение для передвижения растворов по тканям (кровообращение, восходящий и нисходящий токи в растениях). Многим мелким организмам поверхностное натяжение позволяет удерживаться на воде или скользить по ее поверхности.
9. Вода обеспечивает передвижение веществ в клетке и организме, поглощение веществ и выведение продуктов метаболизма.
10. У растений вода определяет тургор клеток, а у некоторых животных выполняет опорные функции, являясь гидростатическим скелетом (круглые и кольчатые черви, иглокожие).
11. Вода - составная часть смазывающих жидкостей (синовиальной - в суставах позвоночных, плевральной - в плевральной полости, перикардиальной - в околосердечной сумке) и слизей (облегчают передвижение веществ по кишечнику, создают влажную среду на слизистых оболочках дыхательных путей). Она входит в состав слюны, желчи, слез и др.
Свойства, функции и значение воды
Минеральные соли . Молекулы солей в водном растворе распадаются на катионы и анионы. Наибольшее значение имеют катионы (К+, Na+, Са2+, Mg2+, NH4+) и анионы (Сl- , Н2Р04 -, НР042- , НС03 -, NO3 2-, SO4 2-) .Некоторые ионы участвуют в активации ферментов, создании осмотического давления в клетке, в процессах мышечного сокращения, свертывании крови и др. Ряд катионов и анионов необходим для синтеза важных органических веществ (например, фосфолипидов, АТФ, нуклеотидов, гемоглобина, хлорофилла и др.), а также аминокислот, являясь источниками атомов азота и серы. Соляная кислота входит в состав желудочного сока. Соли кальция и фосфора присутствуют в костной ткани животных и человека.
Органические вещества. Основой всех органических соединений является углерод (С), который образует связи с другими атомами и их группами. В результате образуются сложные химические соединения, разные по строению и функциям, - макромолекулы (от греч. macros – большой).
Макромолекулы состоят из повторяющихся низкомолекулярных соединений, - мономеров (от греч. monos – один).
Полимер (от греч. poly – много) – макромолекула, образованная мономерами.
В молекулах полимеров мономеры могут быть одинаковые или разные. В зависимости от того, какие мономеры входят в состав полимеров, полимеры делятся на следующие группы:
Полимеры
Регулярные Нерегулярные
А-А-А-А-A-A- - А-В-А-С- В-А-А-D- C- A-
A-S-D-A-S-D-A-S-D-
Полимеры, входящие в состав живых организмов, называются биополимеры, свойства которых зависят от строения их молекул, числа и разнообразия мономеров. Биополимеры универсальны, так как построены по единому плану у всех живых организмов. Разнообразие свойств биополимеров обусловлено различным сочетанием мономеров, образующих различные варианты. Свойства биополимеров проявляются только в живой клетке.
Углеводы, или сахариды , - органические соединения, в состав которых входят углерод, водород и кислород. Название «углеводы» они получили из-за своего химического состава: общая формула большинства из них Сn(H2O)n.
Состав и строение углеводов
Моносахариды – простые сахара, имеющие общую формулу (СН2О)n , где n=3-9. Среди моносахаридов различают триозы (3С), тетраозы (4С), пентозы (5С) – рибоза, дезоксирибоза, гексозы (6С) – глюкоза, галактоза. Моносахариды хорошо растворяются в воде, они сладкие на вкус. Фруктоза входит в состав меда, находится в плодах, зеленых частях растений. Глюкоза находится в плодах, крови, лимфе, является основным источником энергии, входит в состав дисахаридов и полисахаридов.
Дисахариды – вещества, образованные в результате конденсации двух молекул моносахаридов с потерей одной молекулы воды. У растений - это сахароза (С12Н22О11) и мальтоза, у животных – лактоза. Сахароза – основная транспортная форма углеводов в растениях. Лактоза образуется в молочной железе и присутствует в молоке.
глюкоза + глюкоза = мальтоза;
глюкоза + галактоза = лактоза;
глюкоза + фруктоза = саxароза.
По своим свойствам дисахариды близки к моносахаридам. Они хорошо растворяются в воде и имеют сладкий вкус.
Полисахариды – это высокомолекулярные углеводы, образованные путем соединения большого числа молекул моносахаридов, У растений – крахмал, целлюлоза (клетчатка), формула (С6Н10О5)n ; у животных – гликоген, хитин. Целлюлоза – основной опорный компонент клеточной стенки у растений. Крахмал – основной резервный углевод растений. Гликоген – резервный полисахарид животных (накапливается в печени и мышцах. Хитин входит в состав покровов членистоногих, обеспечивает прочность покровных структур грибов.
Локализация в клетке и организме: клеточная стенка, клеточные включения, клеточный сок растений, покровы членистоногих.
Функции углеводов :
1) Энергетическая. Углеводы – это основной источник энергии для организмов. В процессе окисления 1 г углеводов освобождается 17,6 кДж.
2) Структурная. Клеточные стенки растений построены из целлюлозы. Покровы тела членистоногих, клеточные стенки грибов состоят из хитина. Углеводы входят в состав органоидов, молекул ДНК и РНК.
3) Запасающая. Эту функцию выполняют у растений крахмал, у животных гликоген. Они обладают способностью накапливаться в клетках и расходоваться по мере возникновения потребности в энергии.
4) Защитная. Железы выделяют секреты, которые содержат углеводы. Секреты защищают стенки полых органов (желудок, кишечник) от механических повреждений, проникновения болезнетворных бактерий.
Липиды - это жироподобные вещества, большинство из которых состоит из жирных кислот и трехатомного спирта; это сложные эфиры высших жирных кислот и трехатомного спирта глицерина.
Жиры – наиболее простые и широко распространенные липиды. Жидкие жиры называются маслами. У животных масла встречаются в молоке, но чаще встречаются у растений в семенах, плодах.
Состав и строение липидов
Место синтеза в клетке: на мембранах гладкой эндоплазматической сети.
Локализация в клетке и организме: клеточная мембрана, клеточные включения, подкожная жировая клетчатка и сальники.
Функции липидов :
1) Энергетическая. Липиды – «энергетическое депо». При окислении 1 г липидов до СО2 и Н2О освобождается 38,9 кДж, что в два раза больше по сравнению с углеводами и белками.
2) Структурная. Липиды принимают участие в построении мембран клеток и образовании важных биологических соединений, например, гормонов, витаминов.
3) Запасающая. В растениях чаще накапливаются масла, а не жиры. Семена сои и подсолнечника богаты маслами.
4) Защитная и теплоизоляционная. Жиры плохо проводят тепло. Они откладываются под кожей животных, у некоторых достигают такие скопления толщины до 1 м, например, у китов. Жировой слой защищает животных от переохлаждения. Жировая ткань выполняет функцию терморегулятора. У китов, кроме того, он играет еще и другую роль - способствует плавучести. Благодаря низкой теплопроводности слой подкожного жира помогает сохранить тепло, что позволяет, например, многим животным обитать в условиях холодного климата.
5) Смазывающая и водоотталкивающая. Воск покрывает кожу, шерсть, перья, делает их более эластичными и предохраняет от влаги. Восковой налет имеют листья и плоды многих растений. Такой слой защищает листья во время сильных дождей от намокания.
6) Регуляторная. Многие биологически активные вещества (половые гормоны - тестостерон у
мужчин и прогестерон у женщин), витамины (A, D, E) являются соединениями липидной
7) Источник метаболической воды. Одним из продуктов окисления жира является вода, которая
очень важна для некоторых обитателей животного мира пустынь, например, для верблюдов.
Жир, который запасают эти животные в горбах, является источником воды. Окисление 100 г
жира дает примерно 105 г воды. Необходимую для жизнедеятельности воду медведи, сурки и
другие животные, впадающие в спячку, получают в результате окисления жира.
8) В миелиновых оболочках аксонов нервных клеток липиды являются изоляторами при проведении нервных импульсов.
9) Воск используется пчелами в строительстве сот.
Липиды могут образовывать комплексы с другими биологическими молекулами - белками и сахарами.
Белки, или протеины (от греч. protos – первый) – самые многочисленные, разнообразные и имеющие первостепенное значение органические соединения. Белки – макромолекулы, так как имеют большие размеры.
Химический состав молекул белка: углерод, кислород, водород, азот, сера, также могут быть фосфор, железо, цинк, медь.
Белки - это полимеры, состоящие из повторяющихся низкомолекулярных мономеров. Аминокислоты – мономеры белковых молекул. Известно около 200 аминокислот, встречающихся в живых организмах, но только 20 из них входят в состав белков. Это так называемые основные, или белокобразующие аминокислоты. 20 аминокислот обеспечивают многообразие белков. У растений все необходимые аминокислоты синтезируются из первичных продуктов фотосинтеза. Человек и животные не способны синтезировать ряд аминокислот и должны получать их в готовом виде вместе с пищей. Такие аминокислоты называются незаменимыми. К ним относятся лизин, валин, лейцин, изолейцин, треонин, фенилаланин, триптофан, метионин, аргинин и гистидин (всего 10).
Строение аминокислоты:
Между аминогруппой одной аминокислоты и карбоксильной группой другой аминокислоты образуется ковалентная связь, которая называется пептидная связь, а молекула белка – полипептид .
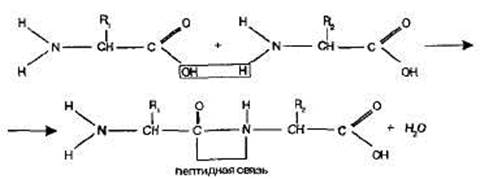
В растворе аминокислоты могут выступать в роли как кислот, так и оснований, т. е. они являются амфотерными соединениями. Карбоксильная группа -СООН способна отдавать протон, функционируя как кислота, а аминная - NH2 - принимать протон, проявляя таким образом свойства основания.
Структура белков. Каждому белку в определенной среде свойственна особая пространственная структура. При характеристике пространственной (трехмерной) структуры выделяют четыре уровня организации молекул белков.
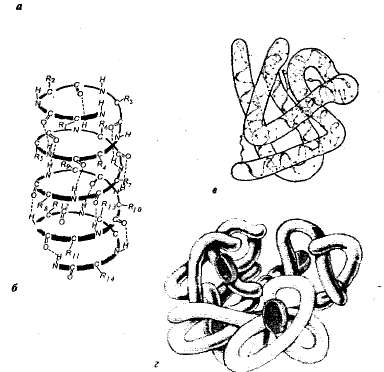
Уровни структурной организации белка: а - первичная структура - аминокислотная последовательность белка; б - вторичная структура - полипептидная цепь закручена в виде спирали; в - третичная структура белка; г - четвертичная структура гемоглобина.
Место синтеза белков в клетке: на рибосомах.
Локализация белков в клетке и организме: присутствуют во всех органоидах и цитоплазматическом матриксе.
Пространственная структура белка:
Первичная структура белка – последовательность аминокислот, соединенных друг с другом пептидными связями в полипептидную цепь. От первичной структуры зависят все свойства и функции белков. Замена одной-единственной аминокислоты в составе молекул белка или нарушение порядка в их расположении обычно влечет за собой изменение функции белка.
Вторичная структура белковой молекулы достигается ее спирализацией: полипептидная цепь, состоящая из последовательно соединенных аминокислот, закручивается в спираль, образуются непрочные водородные связи между – СО- и – NН- группами.
При образовании третичной структуры спирализованная белковая молекула еще многократно сворачивается, образуя шарик – глобулу. Прочность третичной структуры определяется различными связями, например, дисульфидными связями (-S-S-), ионные, водородные, гидрофобное взаимодействие.
Четвертичная структура - это соединение, состоящее из нескольких молекул белка, имеющих третичную структуру. Химические связи - ионные, водородные, гидрофобное взаимодействие.
И так, первичная структура – это линейная структура, в виде полипептидной цепи; вторичная – спиральная, за счет водородных связей; третичная – глобулярная; четвертичная – объединение нескольких молекул белка с третичной структурой.
Свойство белка – денатурация - нарушение природной структуры белка, которая бывает обратимая, если не разрушена первичная структура, и необратимая, если первичная структура разрушена.
Воздействие факторов среды
(температура, химические вещества, излучение и др.)
Денатурация белка (разрушение структур)
Ренатурация – полное восстановление структуры белка.
Под влиянием различных химических и физических факторов (обработка спиртом, ацетоном, кислотами, щелочами, высокой температурой, облучением, высоким давлением и т. д.) происходит изменение вторичной, третичной и четвертичной структур белка вследствие разрыва водородных и ионных связей. Процесс нарушения естественной структуры белка называется денатурацией. При этом наблюдается уменьшение растворимости белка, изменение формы и размеров молекул, потеря ферментативной активности и т. д. Процесс денатурации может быть полным или частичным. В некоторых случаях переход к нормальным условиям среды сопровождается самопроизвольным восстановлением естественной структуры белка. Такой процесс называется ренатурацией.
Простые и сложные белки. По химическому составу выделяют белки простые и сложные. К простым относятся белки, состоящие только из аминокислот, а к сложный - белки, содержащие белковую часть и небелковую - ионы металлов, остаток фосфорной кислоты, углеводы, липиды и др.
Функции белков :
1) Ферментативная , или каталитическая. Катализаторы – это вещества, ускоряющие химические реакции. Ферменты – это катализаторы биохимических реакций. Ферменты ускоряют реакции в организме в десятки и сотни тысяч раз. Они высокоспецифичны, так как каждый фермент катализирует только определенную реакцию.
Ферменты = Биокатализаторы (ускорители химических реакций, протекающих в клетках)
2) Структурная. Белки входят в состав всех мембран и органоидов клетки (например, в соединении с РНК белок образует рибосомы).
3) Энергетическая . При распаде 1 г белков до конечных продуктов (СО2, Н2О и азотсодержащие вещества) выделяется 17,6 кДж.
4) Запасающая. Эту функцию выполняют белки – источники питания (белок яйца – альбумин,
белок молока – казеин, клетки эндосперма и яйцеклетки).
5) Защитная. Все живые клетки и организмы имеют защитные системы. У человека и животных - это иммунная защита. В лимфоцитах образуются антитела – защитные белки, которые обезвреживают чужеродные тела. Другой пример защитной функции – свертывание белка фибриногена в крови, что приводит к образованию сгустка крови – тромба, который закупоривает сосуд, кровотечение прекращается. Механическую защиту обеспечивают роговые образования – волосы, рога, копыта. В состав этих образований входят белки. Растения тоже образуют защитные белки, например, алкалоиды, благодаря которым покровы растений становятся более прочными и устойчивыми.
6) Регуляторная. Многие белки – гормоны , регулирующие физиологические процессы (белковую природу имеют инсулин и глюкагон). Клетки поджелудочной железы вырабатывают гормон инсулин, регулирующий содержание глюкозы в крови.
Поджелудочная железа
Гормон инсулин
Глюкоза (в крови) à Гликоген (в клетках печени)
7) Транспортная. Функция транспортных белков заключается в присоединении химических элементов или биологически активных веществ и переносе их к тканям и органам.
Гемоглобин (находится в эритроцитах)
Гемоглобин + кислород Гемоглобин + углекислый газ
8) Двигательная. Сократительные белки участвуют во всех видах движения, к которым способны клетки и организмы. Примеры: движение жгутиков и ресничек у простейших одноклеточных животных, сокращение мышц у многоклеточных животных (белки миозин и актин обеспечивают сокращение мышечных клеток), движение листьев у растений.
9) Сигнальная. Белки, встроенные в мембрану клетки, осуществляют прием сигналов из
внешней среды и передачу информации в клетку. Такие белковые молекулы способны
изменять свою третичную структуру в ответ на действия факторов внешней среды.
10) Токсическая (токсины, обеспечивающие защиту от врагов и умерщвление добычи).
| Функции белка | Характеристика |
| 1. Структурная | Белки входят в состав клеточных мембран и органоидов |
| 2. Энергетическая | При окислении 1 г белков выделяется 17,6 кДж |
| 3. Запасающая | Белки – запасной питательный и энергетический материал |
| 4. Каталитическая, ферментативная | Белки – ферменты, ускоряющие химические реакции |
| 5. Регуляторная | Многие белки – гормоны, регулирующие физиологические процессы |
| 6. Транспортная | Перенос различных веществ (гемоглобин + кислород) |
| 7. Двигательная | Сократительные белки обеспечивают движение (хромосомы к полюсам клетки) |
| 8. Защитная | Защищают организм от чужеродных тел |
| 9. Сигнальная | Осуществляют прием сигналов из внешней среды и передачу информации в клетку |
| 10. Токсическая | Токсины обеспечивают защиту от врагов и умерщвление добычи |
Белки используются как источник энергии редко, поскольку они выполняют ряд других важных функций. Белки обычно используются, когда истощаются такие источники, как углеводы и жиры. Углеводы и жиры откладываются в запас; когда в пище не хватает какого-либо органического соединения, возможно превращение в организме одних органических соединений в другие: белков в жиры и углеводы, углеводы и жиры друг в друга. Но углеводы и жиры не могут превращаться в белки.
УГЛЕВОДЫ ЖИРЫ
Аминокислоты, которые образуются при расщеплении белковых молекул, необходимы для построения новых белков. Недостаток белков в пище является невосполнимым, так как они образуются только из аминокислот. Поэтому белковое голодание опасно для организма.
Нуклеиновые кислоты. АТФ
Нуклеиновые кислоты (от лат. nucleus – ядро) – кислоты, впервые обнаруженные при исследовании ядер лейкоцитов; были открыты в 1868 г. И.Ф. Мишером, швейцарским биохимиком. Биологическое значение нуклеиновых кислот - хранение и передача наследственной информации; они необходимы для поддержания жизни и для ее воспроизведения.
Нуклеиновые кислоты
Нуклеотид ДНК и нуклеотид РНК имеют черты сходства и различия.
Строение нуклеотида ДНК
Строение нуклеотида РНК
Молекула ДНК – двойная цепь, закрученная по спирали.
Молекула РНК представляет собой одиночную нить нуклеотидов, схожую по строению с отдельной нитью ДНК. Только вместо дезоксирибозы РНК включает другой углевод – рибозу (отсюда и название), а вместо тимина – урацил.
Две нити ДНК соединены друг с другом водородными связями. При этом наблюдается важная закономерность: напротив азотистого основания аденин А в одной цепи располагается азотистое основание тимин Т в другой цепи, а против гуанина Г всегда расположен цитозин Ц. Эти пары оснований называют комплементарными парами.
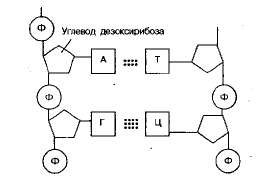
Таким образом, принцип комплементарности (от лат. complementum – дополнение) состоит в том, что каждому азотистому основанию, входящему в нуклеотид, соответствует другое азотистое основание. Возникают строго определенные пары оснований (А – Т, Г – Ц), эти пары специфичны. Между гуанином и цитозином – три водородные связи, а между аденином и тимином возникают две водородные связи в нуклеотиде ДНК, а в РНК две водородные связи возникают между аденином и урацилом.
Водородные связи между азотистыми основаниями нуклеотидов
Г ≡ Ц Г ≡ Ц
В результате у всякого организма число адениловых нуклеотидов равно числу тимидиловых, а число гуаниловых - числу цитидиловых. Благодаря этому свойству последовательность нуклеотидов в одной цепи определяет их последовательность в другой. Такая способность к избирательному соединению нуклеотидов называется комплементарностью, и это свойство лежит в основе образования новых молекул ДНК на базе исходной молекулы (репликации, т. е. удвоения).
Таким образом, количественное содержание азотистых оснований в ДНК подчинено некоторым правилам:
1) Сумма аденина и гуанина равна сумме цитозина и тимина А + Г = Ц + Т.
2) Сумма аденина и цитозина равна сумме гуанина и тимина А + Ц = Г + Т.
3) Количество аденина равно количеству тимина, количество гуанина равно количеству цитозина А = Т; Г = Ц.
При изменении условий ДНК, подобно белкам, может подвергаться денатурации, которая называется плавлением.
ДНК обладает уникальными свойствами: способностью к самоудвоению (репликация, редупликация) и способностью к самовосстановлению (репарация). Репликация обеспечивает точное воспроизведение в дочерних молекулах той информации, которая была записана в материнской молекуле. Но в процессе репликации иногда возникают ошибки. Способность молекулы ДНК исправлять ошибки, возникающие в ее цепях, то есть восстанавливать правильную последовательность нуклеотидов, называется репарацией .
Молекулы ДНК находятся в основном в ядрах клеток и в небольшом количестве в митохондриях и пластидах – хлоропластах. Молекулы ДНК – носители наследственной информации.
Строение, функции и локализация в клетке. Различают три вида РНК. Названия связаны с выполняемыми функциями:
Сравнительная характеристика нуклеиновых кислот
Аденозинфосфорные кислоты - аденозинтрифосфорная кислота (АТФ), аденозиндифосфорная кислота (АДФ), аденозинмонофосфорная кислота (АМФ).
В цитоплазме каждой клетки, а также в митохондриях, хлоропластах и ядрах содержится аденозинтрифосфорная кислота (АТФ). Она поставляет энергию для большинства реакций, происходящих в клетке. С помощью АТФ клетка синтезирует новые молекулы белков, углеводов, жиров, осуществляет активный транспорт веществ, биение жгутиков и ресничек.
АТФпо строению сходна с адениновым нуклеотидом, входящим в состав РНК, только вместо одной фосфорной кислоты в состав АТФ входят три остатка фосфорной кислоты.
Строение молекулы АТФ:
Неустойчивые химические связи, которыми соединены молекулы фосфорной кислоты в АТФ, очень богаты энергией. При разрыве этих связей выделяется энергия, которая используется каждой клеткой для обеспечения процессов жизнедеятельности:
АТФ АДФ + Ф + Е
АДФ АМФ + Ф + Е,
где Ф – фосфорная кислота Н3РО4, Е – освобождающаяся энергия.
Химические связи в АТФ между остатками фосфорной кислоты, богатые энергией, называются макроэргическими связями . Отщепление одной молекулы фосфорной кислоты сопровождается выделением энергии – 40 кДж.
АТФ образуется из АДФ и неорганического фосфата за счет энергии, освобождающейся при окислении органических веществ и в процессе фотосинтеза. Этот процесс называется фосфорилированием.
При этом должно быть затрачено не менее 40 кДж/моль энергии, которая аккумулируется в макроэргических связях. Следовательно, основное значение процессов дыхания и фотосинтеза определяется тем, что они поставляют энергию для синтеза АТФ, с участием которой в клетке выполняется большая часть работы.
АТФ чрезвычайно быстро обновляется. У человека, например, каждая молекула АТФ расщепляется и вновь восстанавливается 2 400 раз в сутки, так что ее средняя продолжительность жизни менее 1 мин. Синтез АТФ осуществляется главным образом в митохондриях и хлоропластах (частично в цитоплазме). Образовавшаяся здесь АТФ направляется в те участки клетки, где возникает потребность в энергии.
АТФ играет важную роль в биоэнергетике клетки: выполняет одну из важнейших функций – накопителя энергии, это универсальный биологический аккумулятор энергии.
Химический состав клетки
В клетках обнаружено около 60 элементов периодической системы Менделеева, встречающихся и в неживой природе. Это одно из доказательств общности живой и неживой природы. В живых организмах наиболее распространены водород, кислород, углерод и азот, которые составляют около 98% массы клеток. Такое обусловлено особенностями химических свойств водорода, кислорода, углерода и азота, вследствие чего они оказались наиболее подходящими для образования молекул, выполняющих биологические функции. Эти четыре элемента способны образовывать очень прочные ковалентные связи посредством спаривания электронов, принадлежащих двум атомам. Ковалентно связанные атомы углерода могут формировать каркасы бесчисленного множества различных органических молекул. Поскольку атомы углерода легко образуют ковалентные связи с кислородом, водородом, азотом, а также с серой, органические молекулы достигают исключительной сложности и разнообразия строения.
Кроме четырех основных элементов в клетке в заметных количествах (10 ые и 100 ые доли процента) содержатся железо, калий, натрий, кальций, магний, хлор, фосфор и сера. Все остальные элементы (цинк, медь, йод, фтор, кобальт, марганец и др.) находятся в клетке в очень малых количествах и поэтому называются микроэлементами.
Химические элементы входят в состав неорганических и органических соединений. К неорганическим соединениям относятся вода, минеральные соли, диоксид углерода, кислоты и основания. Органические соединения – это белки, нуклеиновые кислоты, углеводы, жиры (липиды) и липоиды. Кроме кислорода, водорода, углерода и азота в их состав могут входить другие элементы. Некоторые белки содержат серу. Составной частью нуклеиновых кислот является фосфор. Молекула гемоглобина включает железо, магний участвует в построении молекулы хлорофилла. Микроэлементы, несмотря на крайне низкое содержание в живых организмах, играют важную роль в процессах жизнедеятельности. Йод входит в состав гормона щитовидной железы – тироксина, кобальт – в состав витамина В 12 гормон островковой части поджелудочной железы – инсулин – содержит цинк. У некоторых рыб место железа в молекулах пигментов, переносящих кислород, занимает медь.
Неорганические вещества
Вода. Н 2 О – самое распространенное соединение в живых организмах. Содержание ее в разных клетках колеблется в довольно широких пределах: от 10% в эмали зубов до 98% в теле медузы, но среднем она составляет около 80% массы тела. Исключительно важная роль воды в обеспечении процессов жизнедеятельности обусловлена ее физико-химическими свойствами. Полярность молекул и способность образовывать водородные связи делают воду хорошим растворителем для огромного количества веществ. Большинство химических реакций, протекающих в клетке, может происходить только в водном растворе. Вода участвует и во многих химических превращениях.
Общее число водородных связей между молекулами воды изменяется в зависимости от t°. При t° таяния льда разрушается примерно 15% водородных связей, при t° 40°С – половина. При переходе в газообразное состояние разрушаются все водородные связи. Этим объясняется высокая удельная теплоемкость воды. При изменении t° внешней среды вода поглощает или выделяет теплоту вследствие разрыва или новообразования водородных связей. Таким путем колебания t° внутри клетки оказываются меньшими, чем в окружающей среде. Высокая теплота испарения лежит в основе эффективного механизма теплоотдачи у растений и животных.
Вода как растворитель принимает участие в явлениях осмоса, играющего важную роль в жизнедеятельности клетки организма. Осмосом называют проникновение молекул растворителя через полупроницаемую мембрану в раствор какого-либо вещества. Полупроницаемыми называются мембраны, которые пропускают молекулы растворителя, но не пропускают молекулы (или ионы) растворенного вещества. Следовательно, осмос – односторонняя диффузия молекул воды в направлении раствора.
Минеральные соли. Большая часть неорганических в-в клетки находится в виде солей в диссоциированном, либо в твердом состоянии. Концентрация катионов и анионов в клетке и в окружающей ее среде неодинакова. В клетке содержится довольно много К и очень много Nа. Во внеклеточной среде, например в плазме крови, в морской воде, наоборот, много натрия и мало калия. Раздражимость клетки зависит от соотношения концентраций ионов Na + , K + , Ca 2+ , Mg 2+ . В тканях многоклеточных животных К входит в состав многоклеточного вещества, обеспечивающего сцепленность клеток и упорядоченное их расположение. От концентрации солей в большой мере зависят осмотическое давление в клетке и ее буферные свойства. Буферностью называется способность клетки поддерживать слабощелочную реакцию ее содержимого на постоянном уровне. Буферность внутри клетки обеспечивается главным образом ионами Н 2 РО 4 и НРО 4 2- . Во внеклеточных жидкостях и в крови роль буфера играют Н 2 СО 3 и НСО 3 - . Анионы связывают ионы Н и гидроксид-ионы (ОН -), благодаря чему реакция внутри клетки внеклеточных жидкостей практически не меняется. Нерастворимые минеральные соли (например, фосфорнокислый Са) обеспечивает прочность костной ткани позвоночных и раковин моллюсков.
11. Химический состав клетки. Неорганические вещества клетки.
Органические вещества клетки
Белки. Среди органических веществ клетки белки стоят на первом месте как по количеству (10 – 12% от общей массы клетки), так и по значению. Белки представляют собой высокомолекулярные полимеры (с молекулярной массой от 6000 до 1 млн. и выше), мономерами которых являются аминокислоты. Живыми организмами используется 20 аминокислот, хотя их существует значительно больше. В состав любой аминокислоты входит аминогруппа (-NH 2), обладающая основными свойствами, и карбоксильная группа (-СООН), имеющая кислотные свойства. Две аминокислоты соединяются в одну молекулу путем установления связи HN-CO с выделением молекулы воды. Связь между аминогруппой одной аминокислоты и карбоксилом другой называется пептидной. Белки представляют собой полипептиды, содержащие десятки и сотни аминокислот. Молекулы различных белков отличаются друг от друга молекулярной массой, числом, составом аминокислот и последовательностью расположения их в полипептидной цепи. Понятно поэтому, что белки отличаются огромным разнообразием, их количество у всех видов живых организмов оценивается числом 10 10 – 10 12 .
Цепь аминокислотных звеньев, соединенных ковалентное пептидными связями в определенной последовательности, называется первичной структурой белка. В клетках белки имеют вид спирально закрученных волокон или шариков (глобул). Это объясняется тем, что в природном белке полипептидная цепочка уложена строго определенным образом в зависимости от химического строения входящих в ее состав аминокислот.
Вначале полипептидная цепь сворачивается в спираль. Между атомами соседних витков возникает притяжение и образуются водородные связи, в частности, между NH- и СО- группами, расположенными на соседних витках. Цепочка аминокислот, закрученная в виде спирали, образует вторичную структуру белка. В результате дальнейшей укладки спирали возникает специфичная для каждого белка конфигурация, называемая третичной структурой. Третичная структура обусловлена действием сил сцепления между гидрофобными радикалами, имеющимися у некоторых аминокислот, и ковалентными связями между SH- группами аминокислоты цистеина (S-S- связи). Количество аминокислот гидрофобными радикалами и цистеина, а также порядок их расположения в полипептидной цепочке специфичны для каждого белка. Следовательно, особенности третичной структуры белка определяются его первичной структурой. Биологическую активность белок проявляет только в виде третичной структуры. Поэтому замена даже одной аминокислоты в полипептидной цепочке может привести к изменению конфигурации белка и к снижению или утрате его биологической активности.
В некоторых случаях белковые молекулы объединяются друг с другом и могут выполнять свою функцию только в виде комплексов. Так, гемоглобин – это комплекс из четырех молекул и только в такой форме способен присоединять и транспортировать О. подобные агрегаты представляют собой четвертичную структуру белка. По своему составу белки делятся на два основных класса – простые и сложные. Простые белки состоят только из аминокислот нуклеиновые кислоты (нуклеотиды), липиды (липопротеиды), Ме (металлопротеиды), Р (фосфопротеиды).
Функции белков в клетке чрезвычайно многообразны. Одна из важнейших – строительная функция: белки участвуют в образовании всех клеточных мембран и органоидов клетки, а также внутриклеточных структур. Исключительно важное значение имеет ферментативная (каталитическая) роль белков. Ферменты ускоряют химические реакции, протекающие в клетке, в 10 ки и 100 ни миллионов раз. Двигательная функция обеспечивается специальными сократительными белками. Эти белки участвуют во всех видах движений, к которым способны клетки и организмы: мерцание ресничек и биение жгутиков у простейших, сокращение мышц у животных, движение листьев у растений и др. Транспортная функция белков заключается в присоединении химических элементов (например, гемоглобин присоединяет О) или биологически активных веществ (гормонов) и переносе их к тканям и органам тела. Защитная функция выражается в форме выработки особых белков, называемых антителами, в ответ на проникновение в организм чужеродных белков или клеток. Антитела связывают и обезвреживают чужеродные вещества. Белки играют немаловажную роль как источники энергии. При полном расщеплении 1г. белков выделяется 17,6 кДж (~4,2 ккал).
Углеводы. Углеводы, или сахариды – органические вещества с общей формулой (СН 2 О) n . У большинства углеводов число атомов Н вдвое больше числа атомов О, как в молекулах воды. Поэтому эти вещества и были названы углеводами. В живой клетке углеводы находятся в количествах, не превышающих 1-2, иногда 5% (в печени, в мышцах). Наиболее богаты углеводами растительные клетки, где их содержание достигает в некоторых случаях 90% от массы сухого вещества (семена, клубни картофеля и т.д.).
Углеводы бывают простые и сложные. Простые углеводы называются моносахаридами. В зависимости от числа атомов углевода в молекуле моносахариды называются триозами, тетрозами, пентозами или гексозами. Из шести углеродных моносахаридов – гексоз – наиболее важное значение имеют глюкоза, фруктоза и галактоза. Глюкоза содержится в крови (0,1-0,12%). Пентозы рибоза и дезоксирибоза входят в состав нуклеиновых кислот и АТФ. Если в одной молекуле объединяются два моносахарида, такое соединение называется дисахаридом. Пищевой сахар, получаемый из тростника или сахарной свеклы, состоит из одной молекулы глюкозы и одной молекулы фруктозы, молочный сахар – из глюкозы и галактозы.
Сложные углеводы, образованные многими моносахаридами, называются полисахаридами. Мономером таких полисахаридов, как крахмал, гликоген, целлюлоза, является глюкоза. Углеводы выполняют две основные функции: строительную и энергетическую. Целлюлоза образует стенки растительных клеток. Сложный полисахарид хитин служит главным структурным компонентом наружного скелета членистоногих. Строительную функцию хитин выполняет и у грибов. Углеводы играют роль основного источника энергии в клетке. В процессе окисления 1 г. углеводов освобождается 17,6 кДж (~4,2 ккал). Крахмал у растений и гликоген у животных откладываются в клетках и служат энергетическим резервом.
Нуклеиновые кислоты. Значение нуклеиновых кислот в клетке очень велико. Особенности их химического строения обеспечивают возможность хранения, переноса и передачи по наследству дочерним клеткам информации о структуре белковых молекул, которые синтезируются в каждой ткани на определенном этапе индивидуального развития. Поскольку большинство свойств и признаков клеток обусловлено белками, то понятно, что стабильность нуклеиновых кислот – важнейшее условие нормальной жизнедеятельности клеток и целых организмов. Любые изменения структуры клеток или активности физиологических процессов в них, влияя, таким образом, на жизнедеятельность. Изучение структуры нуклеиновых кислот имеет исключительно важное значение для понимания наследования признаков у организмов и закономерностей функционирования, как отдельных клеток, так и клеточных систем – тканей и органов.
Существуют 2 типа нуклеиновых кислот – ДНК и РНК. ДНК – полимер, состоящий из двух нуклеотидных спиралей, заключенных так, что образуется двойная спираль. Мономеры молекул ДНК представляют собой нуклеотиды, состоящие из азотистого основания (аденина, тимина, гуанина или цитозина), углевода (дезоксирибозы) и остатка фосфорной кислоты. Азотистые основания в молекуле ДНК соединены между собой неодинаковым количеством Н-связей и располагаются попарно: аденин (А) всегда против тимина (Т), гуанин (Г) против цитозина (Ц). Схематически расположение нуклеотидов в молекуле ДНК можно изобразить так:
Рис.1.Расположение нуклеотидов в молекуле ДНК
Из рис.1. видно, что нуклеотиды соединены друг с другом не случайно, а избирательно. Способность к избирательному взаимодействию аденина с тимином и гуанина с цитозином называется комплементарностью. Комплементарное взаимодействие определенных нуклеотидов объясняется особенностями пространственного расположения атомов в их молекулах, которые позволяют им сближаться и образовывать Н-связи. В полинуклеотидной цепочке соседние нуклеотиды связаны между собой через сахар (дезоксирибозу) и остаток фосфорной кислоты. РНК так же, как и ДНК, представляет собой полимер, мономерами которого являются нуклеотиды. Азотистые основания трех нуклеотидов те же самые, что входят в состав ДНК (А, Г, Ц); четвертое – урацил (У) – присутствует в молекуле РНК вместо тимина. Нуклеотиды РНК отличаются от нуклеотидов ДНК и по строению входящего в их состав углевода (рибоза вместо дизоксирибозы).
В цепочке РНК нуклеотиды соединяются путем образования ковалентных связей между рибозой одного нуклеотида и остатком фосфорной кислоты другого. По структуре различаются двух цепочечные РНК. Двух цепочечные РНК являются хранителями генетической информации у ряда вирусов, т.е. выполняют у них функции хромосом. Одно цепочечные РНК осуществляют перенос информации о структуре белков от хромосомы к месту их синтеза и участвуют в синтезе белков.
Существует несколько видов одно цепочечной РНК. Их названия обусловлены выполняемой функцией или местом нахождения в клетке. Большую часть РНК цитоплазмы (до 80-90%) составляет рибосомальная РНК (рРНК), содержащаяся в рибосомах. Молекулы рРНК относительно невелики и состоят в среднем из 10 нуклеотидов. Другой вид РНК (иРНК), переносящие к рибосомам информацию о последовательности аминокислот в белках, которые должны синтезироваться. Размер этих РНК зависит от длины участка ДНК, на котором они были синтезированы. Транспортные РНК выполняют несколько функций. Они доставляют аминокислоты к месту синтеза белка, "узнают" (по принципу комплементарности) триплет и РНК, соответствующий переносимой аминокислоте, осуществляют точную ориентацию аминокислоты на рибосоме.
Жиры и липоиды. Жиры представляют собой соединения жирных высокомолекулярных кислот и трехатомного спирта глицерина. Жиры не растворяются в воде – они гидрофобны. В клетке всегда есть и другие сложные гидрофобные жироподобные вещества, называемые липоидами. Одна из основных функций жиров – энергетическая. В ходе расщепления 1 г. жиров до СО 2 и Н 2 О освобождается большое количество энергии – 38,9 кДж (~9,3 ккал). Содержание жира в клетке колеблется в пределах 5-15% от массы сухого вещества. В клетках живой ткани количество жира возрастает до 90%. Главная функция жиров в животном (и отчасти - растительном) мире - запасающая.
При полном окислении 1 г жира (до углекислого газа и воды) выделяется около 9 ккал энергии. (1 ккал = 1000 кал; калория (кал, cal) - внесистемная единица количества работы и энергии, равная количеству теплоты, необходимому для нагревания 1 мл воды на 1 °C при стандартном атмосферном давлении 101,325 кПа; 1 ккал = 4,19 кДж). При окислении (в организме) 1 г белков или углеводов выделяется только около 4 ккал/г. У самых разных водных организмов - от одноклеточных диатомовых водорослей до гигантских акул - жир случит «поплавком», уменьшая среднюю плотность тела. Плотность животных жиров составляет около 0,91-0,95 г/см³. Плотность костной ткани позвоночных близка к 1,7-1.8 г/см³, а средняя плотность большинства других тканей близка к 1 г/см³. Понятно, что жира нужно довольно много, чтобы «уравновесить» тяжелый скелет.
Жиры и липоиды выполняют и строительную функцию: они входят в состав клеточных мембран. Благодаря плохой теплопроводности жир способен к защитной функции. У некоторых животных (тюлени, киты) он откладывается в подкожной жировой ткани, образуя слой толщиной до 1 м. Образование некоторых липоидов предшествует синтезу ряда гормонов. Следовательно, этим веществам присуща и функция регуляции обменных процессов.
12. Деление клетки.
Клетка в своей жизни проходит разные состояния: фазу роста и фазы подготовки к делению и деления. Клеточный цикл – переход от деления к синтезу веществ, составляющих клетку, а затем опять к делению – можно представить на схеме в виде цикла, в котором выделяют несколько фаз.
Описано три способа деления эукариотических клеток: амитоз (прямое деление), митоз (непрямое деление) и мейоз (редукционное деление).
Амитоз – относительно редкий способ деления клетки. При амитозе интерфазное ядро делится путем перетяжки, равномерное распределение наследственного материала не обеспечивается. Нередко ядро делится без последующего разделения цитоплазмы и образуются двухъядерные клетки. Клетка, претерпевшая амитоз, в дальнейшем не способна вступать в нормальный митотический цикл. Поэтому амитоз встречается, как правило, в клетках и тканях, обреченных на гибель.
Митоз. Митоз, или непрямое деление, - основной способ деления эукариотических клеток. Митоз – это деление ядра, которое приводит к образованию двух дочерних ядер, в каждом из которых имеется точно такой же набор хромосом, что и был в родительском ядре. Имеющиеся в клетке хромосомы удваиваются, выстраиваются в клетке, образуя митотическую пластинку, к ним прикреплены нити веретена деления, которые растягиваются к полюсам клетки и клетка делится, образуя две копии исходного набора.
При образовании гамет, т.е. половых клеток – сперматозоидов и яйцеклеток – происходит деление клетки, называемое мейозом. Исходная клетка имеет диплоидный набор хромосом, которые затем удваиваются. Но, если при митозе в каждой хромосоме хроматиды просто расходятся, то при мейозе хромосома (состоящая из двух хроматид) тесно переплетается своими частями с другой, гомологичной ей хромосомой (также состоящей из двух хроматид), и происходит кроссинговер - обмен гомологичными участками хромосом. Затем уже новые хромосомы с перемешанными «мамиными» и «папиными» генами расходятся и образуются клетки с диплоидным набором хромосом, но состав этих хромосом уже отличается от исходного, в них произошла рекомбинация. Завершается первое деление мейоза, и второе деление мейоза происходит без синтеза ДНК, поэтому при этом делении количество ДНК уменьшается вдвое. Из исходных клеток с диплоидным набором хромосом возникают гаметы с гаплоидным набором. Из одной диплоидной клетки образуются четыре гаплоидных клетки. Фазы деления клетки, которые следуют за интерфазой, называются профаза, метафаза, анафаза, телофаза и после деления опять интерфаза.
Профаза – самая длительная фаза митоза, когда происходит перестройка всей структуры ядра для деления. В профазе происходит укорочение и утолщение хромосом вследствие их спирализации. В это время хромосомы двойные (удвоение происходит в S-периоде интерфазы), состоят из двух хроматид, связанных между собой в области первичной перетяжки осбой структурой – цетромерой. Одновременно с утолщением хромосом исчезает ядрышко и фрагментируется (распадается на отдельные цистерны) ядерная оболочка. После распада ядерной оболочки хромосомы свободно и беспорядочно лежат в цитоплазме. Начинается формирование ахромативного веретена – веретена деления, которое представляет систему нитей, идущих от полюсов клетки. Нити веретена имеют диаметр около 25нм. Это пучки микротрубочек, состоящих из субъедениц белка тубулина. Микротрубочки начинают формироваться со стороны центриолей либо со стороны хромосом (в клетках растений).
Метафаза. В метафазе завершается образование веретена деления, которое состоит из микротрубочек двух типов: хромосомных, которые связываются с центромерами хромосом, и ценросомных (полюсных), которые тянутся от полюса к полюсу клетки. Каждая двойная хромосома прикрепляется к микротрубочкам веретена деления. Хромосомы как бы выталкиваются микротрубочками в область экватора клетки, т.е. располагаются на равном расстоянии от полюсов. Они лежат в одной плоскости и образуют так называемую экваториальную, или метафазную пластинку. В метафазе отчетливо видно двойное строение хромосом, соединенных только в области центромеры. Именно в этот период легко подсчитать число хромосом, изучать их морфологические особенности.
Анафаза начинается делением центромеры. Каждая из хроматид одной хромосомы становится самостоятельной хромосомой. Сокращение тянущих нитей ахроматинового веретена увлекает их к противоположным полюсам клетки. В результате у каждого из полюсов клетки оказывается столько же хромосом, сколько было их в материнской клетке, причем набор их одинаков.
Телофаза – последняя фаза митоза. Хромосомы деспирализуются, становятся плохо заметными. На каждом из полюсов вокруг хромосом воссоздается ядерная оболочка. Формируются ядрышки, веретено деления исчезает. В образовавшихся ядрах каждая хромосома состоит теперь всего из одной хроматиды, а не из двух.
Каждое из вновь образовавшихся ядер получило весь объем генетической информации, которым обладала ядерная ДНК материнской клетки. В результате митоза оба дочерних ядра имеют одинаковое количество ДНК и одинаковое число хромосом, такое же, как в материнском.
Цитокинез – после образования в телофазе двух новых ядер происходит деление клетки и формирование в экваториальной плоскости перегородки – клеточной пластинки.
В ранней телофазе между двумя дочерними ядрами, не достигая их, формируется цилиндрическая система волокон, называемая фрагмопластом, которая также как и волокна ахроматинового веретена, состоит из микротрубочек и связаны с ним. В центре фрагмопласта на экваторе между дочерними ядрами скапливаются пузырьки Гольджи, содержащие пектиновые вещества. Они сливаются друг с другом и дают начало клеточной пластинке, а их мембраны участвуют в построении плазмолемм по обеим сторонам пластинки. Клеточная пластинка закладывается в виде диска, взвешенного в фрагмопласте. Волокна фрагмопласта, видимо, контролируют направление движения пузырьков Гольджи. Клеточная пластинка растет центробежно по направлению к стенкам материнской клетки за счет включения в нее полисахаридов все новых и новых пузырьков Гольджи. Клеточная пластинка имеет полужидкую консистенцию, состоит из аморфного протопектина и пектатов магния и кальция. В это время из трубчатого ЭР образуются плазмодесмы. Расширяющийся фрагмопласт постепенно приобретает форму бочонка, позволяя клеточной пластинке расти латерально, пока она не соединится со стенками материнской клетки. Фрагмопласт исчезает, обособление двух дочерних клеток заканчивается. Каждый протопласт откладывает на клеточную пластинку свою первичную клеточную стенку.
Цитокинез с помощью клеточной пластинки происходит у всех высших растений и некоторых водорослей. У остальных организмов клетки делятся внедрением клеточной оболочки, которая постепенно углубляется и разделяет клетки.
Биологическое значение митоза состоит в строго одинаковом распределении между дочерними клетками материальных носителей наследственности – молекул ДНК, входящих в состав хромосом. Благодаря равномерному разделению реплицированных хромосом между дочерними клетками обеспечивается образование генетически равноценных клеток и сохраняется преемственность в ряду клеточных поколений. Это обеспечивает таки важные моменты жизнедеятельности, как эмбриональное развитие и рост организмов, восстановление органов и тканей после повреждения. Митотическое деление клеток является также цитологической основой бесполого размножения организмов.
Мейоз. Мейоз – это особый способ деления клеток, в результате которого происходит редукция (уменьшение) числа хромосом вдвое и переход клеток из диплоидного состояния (2n) в гаплоидное (n). Мейоз – единый, непрерывный процесс состоящий из двух последовательных делений, каждое из которых можно разделть на те же, что и в митозе, четыре фазы: профазу, метафазу, анафазу и телофазу. Обоим делениям предшествует одна интерфаза. В синтетическом периоде интерфазы до начала мейоза удваивается количество ДНК и каждая хромосома становится двухроматидной.
Первое мейотическое, или редукционное, деление.
Профаза I продолжается от нескольких часов до нескольких недель. Хромосомы спирализуются. Гомологичные хромосомы коньюгируют, образуя пары – биваленты. Бивалент состоит из четырех хроматид двух гомологичных хромосом. В бивалентах осуществляется кроссинговер – обмен гомологичными участками гомологичных хромосом, что приводит к их глубокому преобразованию. Во время коссинговера происходит обмен блоками генов, что объясняет генетическое разнообразие потомства. К концу профазы исчезает ядерная оболочка и ядрышко, формируется ахроматиновое веретено.
Метафаза I – биваленты собираются в экваториальной плоскости клетки. Ориентирование материнской и отцовской хромосомы из каждой гомологичной пары к одному или другому полюсу веретена деления является случайным. К центромере каждой из хромосом присоединяется тянущая нить ахроматинового веретена. Две сетринские хроматиды не разделяются.
Анафаза I – происходит сокращение тянущих нитей, и к полюсам расходятся двухроматидные хромосомы. Гомологичные хромосмы каждого из бивалентов уходят к противоположным полюсам. Расходятся случайно перераспределенные гомологичные хромосомы каждой пары (независимое распределение), и на каждом из полюсов собирается половинное число (гаплоидный набор) хромосом, образуется два гаплоидных набора хромосом.
Телофаза I – у полюсов веретена собирается одиночный, гаплоидный, набор хромосом, в котором каждый вид хромосом представлен уже не парой, а одной хромосомой, состоящей из двух хроматид. В короткой по продолжительности телофазе I восстанавливается ядерная оболочка, после чего материнская клетка делится на две дочернии.
Второе мейотическое деление следует сразу же после первого и сходно с обычным митозом (поэтому его часто называют митозом мейоза), только клетки, вступающие в него, несут гаплоидный набор хромосом.
Профаза II – непродолжительная.
Метафаза II – снова образуется веретено деления, хромосомы выстраиваются в экваториальной плоскости и центормерами прикрепляются к микротрубочкам веретена деления.
Анафаза II – осуществляется разделение их ценромер и каждая хроматида становится самостоятельной хромосомой. Отделившиеся друг от друга дочерние хромосомы направляются к полюсам веретена.
Телофаза II – завершается расхождение сестринских хромосом к полюсам и наступает деление клеток: из двух гаплоидных клеток образуются 4 клетки с гаплоидным набором хромосом.
Редукционное деление является как бы регулятором, препятствующим непрерывному увеличению числа хромосом при слиянии гамет. Не будь такого механизма, при половом размножении число хромосом удваивалось бы в каждом новом поколении. Т.е. благодаря мейозу поддерживается определенное и постоянное число хромосом во всех поколениях каждого вида растений, животных, протист и грибов. Другое значение заключается в обеспечении разнообразия генетического состава гамет как в результате кроссинговера, так и в результате различного сочетания отцовских и материнских хромосом при их расхождении в анафазе I мейоза. Это обеспечивает появление разнообразного и разнокачественного потомства при половом размножении организмов.
13. Процесс фотосинтеза в растительной клетке.
Процесс фотосинтеза состоит из двух последовательных и взаимосвязанных этапов: светового (фотохимического) и темнового (метаболического). На первой стадии происходит преобразование поглощенной фотосинтетическими пигментами энергии квантов света в энергию химических связей высокоэнергетического соединения АТФ и универсального восстановителя НАДФН - собственно первичных продуктов фотосинтеза, или так называемой «ассимиляционной силы». В темновых реакциях фотосинтеза происходит использование образовавшихся на свету АТФ и НАДФН в цикле фиксации углекислоты и ее последующего восстановления до углеводов.
У всех фотосинтезирующих организмов фотохимические процессы световой стадии фотосинтеза происходят в особых энергопреобразующих мембранах, называемых тилакоидными, и организованы в так называемую электрон-транспортную цепь. Темновые реакции фотосинтеза осуществляются вне тилакоидных мембран (в цитоплазме у прокариот и в строме хлоропласта у растений). Таким образом, световая и темновая стадии фотосинтеза разделены в пространстве и во времени.
Интенсивность фотосинтеза древесных растений широко варьирует в зависимости от взаимодействия многих внешних и внутренних факторов, причем эти взаимодействия изменяются во времени и различны у разных видов.
Фотосинтетическую способность иногда оценивают по чистому приросту сухой массы. Такие данные имеют особое значение, потому что прирост представляет собой среднее истинное увеличение массы за большой промежуток времени в условиях внешней среды, включающих обычные периодически наступающие стрессы.
Некоторые виды покрытосеменных эффективно осуществляют фотосинтез как при низкой, так и при высокой интенсивности света. Многие голосеменные гораздо более продуктивны при высокой освещенности. Сравнение этих двух групп при низкой и высокой интенсивности света часто дает различное представление о фотосинтетической способности с точки зрения накопления питательных веществ. Кроме того, голосеменные часто накапливают некоторое количество сухой массы в период покоя, тогда как листопадные покрытосеменные теряют ее вследствие дыхания. Поэтому голосеменное растение с несколько более низкой интенсивностью фотосинтеза, чем листопадное покрытосеменное во время периода роста, может накапливать в течение года столько же или даже больше общей сухой массы благодаря гораздо большей продолжительности периода фотосинтетической активности.
14. Основные виды растительных тканей. Покровные ткани растений (кожица).
Ткани – это группы клеток, имеющие однородное строение, одинаковое происхождение и выполняющие одну и ту же функцию.
В зависимости от выполняемой функции различают следующие типы тканей: образовательные (меристемы), основные, проводящие, покровные, механические, выделительные. Клетки, составляющие ткань и имеющие более или менее одинаковое строение и функции, называют простыми, если клетки неодинаковые, то ткань называют сложной или комплексной.
Ткани делят на образовательные, или меристемные, и постоянные (покровные, проводящие, основные и т.д.).
Классификация тканей.
1. Образовательные ткани (меристемы): 1) верхушечные;
2) боковые: а) первичные (прокамбий, перицикл);
б) вторичные (камбий, феллоген)
3) вставочные;
4) раневые.
2. Основные: 1) ассимиляционная паренхима;
2) запасающая паренхима.
3. Проводящие: 1) ксилема (древесина);
2) флоэма (луб).
4. Покровные (пограничные): 1) наружные: а) первичные (эпидерма);
б) вторичные (перидерма);
в) третичные (корка, или ритидом)
2) наружные: а) ризодерма;
б) веламен
3) внутренние: а) эндодерма;
б) экзодерма;
в) обкладочные клетки проводящих пучков в листьях
5. Механические (опорные, скелетные) ткани: 1) колленхима;
2) склеренхима: а) волокна;
б) склереиды
6. Выделительные ткани (секреторные).
2. Образовательные ткани. Образовательные ткани, или меристемы, - это постоянно молодые, активно делящиеся группы клеток. Находятся они в местах роста разных органов: кончиках корней, верхушках стеблей и т.д. Благодаря меристемам происходят рост растения и образование новых постоянных тканей и органов.
В зависимости от местоположения в теле растения образовательная ткань может быть верхушечной, или апикальной, боковой, или латеральной, вставочной, или интеркалярной, и раневой. Образовательные ткани делят на первичные и вторичные. Так, верхушечные меристемы всегда первичные, они определяют рост растения в длину. У низкоорганизованных высших растений (хвощи, некоторые папоротники) верхушечные меристемы слабо выражены и представлены всего лишь одной начальной, или инициальной делящейся клеткой. У голосеменных и покрытосеменных верхушечные меристемы хорошо выражены и представлены многими инициальными клетками, образующими конусы нарастания. Латеральные меристемы, как правило, вторичны и за счет них происходит разрастание осевых органов (стеблей, корней) в толщину. К боковым меристемам относят камбий и пробковый камбий (феллоген), деятельность которого способствует образованию пробки в корнях и стеблях растения, а также особую ткань проветривания – чечевички. Боковая меристема, как и камбий, образует клетки древесины и луба. В неблагоприятные периоды жизни растения деятельность камбия замедляется или совсем прекращается. Интеркалярные, или вставочные, меристемы чаще всего первичны и сохраняются в виде отдельных участков в зонах активного роста, например у основания междоузлий и у основания черешков листьев злаков.
3. Покровные ткани. Покровные ткани защищают растение от неблагоприятных воздействий внешней среды: солнечного перегрева, излишнего испарения, резкого перепада температуры воздуха, иссушающего ветра, механического воздействия, от проникновения вовнутрь растения болезнетворных грибов и бактерий и т.д. Различают первичную и вторичную покровные ткани. К первичным покровным тканям относятся кожица, или эпидерма, и эпиблема, к вторичным – перидерма (пробка, пробковый камбий и феллодерма).
Кожица, или эпидерма, покрывает все органы однолетних растений, молодые зеленые побеги многолетних древесных растений текущего вегетационного периода, надземные травянистые части растений (листья, стебли и цветки). Эпидерма чаще всего состоит из одного слоя плотно сомкнутых клеток без межклеточного пространства. Она легко снимается и представляет собой тонкую прозрачную пленку. Эпидерма – живая ткань, состоит из постепенного слоя протопласта с лейкопластами и ядром, крупной вакуоли, занимающей почти всю клетку. Стенка клеток в основном целлюлозная. Наружная стенка эпидермальных клеток более утолщенная, боковые и внутренние – тонкие. Боковые и внутренние стенки клеток имеют поры. Основная функция эпидермы – регуляция газообмена и транспирации, осуществляемая в основном через устьица. Вода и неорганические вещества проникают через поры.
Клетки эпидермы разных растений неодинаковы по форме и размерам. У многих однодольных растений клетки вытянуты в длину, у большинства двудольных имеют извилистые боковые стенки, что повышает плотность их сцепления друг с другом. Эпидерма верхней и нижней частей листа также отличается своим строением: на нижней стороне листа в эпидерме большее число устьиц, а на верхней стороне их гораздо меньше; на листьях водных растений с плавающими на поверхности листьями (кубышка, кувшинка) устьица есть только на верхней стороне листа, а у полностью погруженных в воду растений устьица отсутствуют.
Устьица – высокоспециализированные образования эпидермы, состоят из двух замыкающих клеток и щелевидного образования между ними – устьичной щели. Замыкающие клетки, имеющие полулунную форму, регулируют размер устьичной щели; щель может открываться и закрываться в зависимости от тургорного давления в замыкающих клетках, содержания диоксида углерода в атмосфере и других факторов. Так, днем, когда клетки устьиц участвуют в фотосинтезе, тургорное давление в устьичных клетках высокое, устьичная щель открыта, ночью, наоборот, закрыта. Подобное явление наблюдается в засушливое время и при увядании листьев, связано с приспособлением устьиц запасать влагу внутри растения. У многих видов, произрастающих в районах с избыточным увлажнением, особенно во влажных тропических лесах, имеются устьица, через которые выделяется вода. Устьица получили название гидатоды. Вода в виде капель выделяется наружу и капает с листьев. «Плач» растения – своеобразный предсказатель погоды и по научному называется гуттацией. Гидатоды расположены по краю листа, у них нет механизма открывания и закрывания.
В эпидерме многих растений есть защитные приспособления от неблагоприятных условий: волоски, кутикула, восковой налет и др.
Волоски (трихомы) – своеобразные выросты эпидермы, они могут покрывать все растение или некоторые его части. Волоски бывают живыми и мертвыми. Волоски способствуют уменьшению испарения влаги, предохраняют растение от перегрева, поедания животными и от резких колебаний температуры. Поэтому волосками чаще всего покрыты растения аридных – засушливых областей, высокогорий, приполярных районов земного шара, а также растения засоренных местообитаний.
Волоски бывают одноклеточными и многоклеточными. Одноклеточные волоски представлены в виде сосочков. Сососчки встречаются на лепестках многих цветков, придавая им бархатистость (тагетисы, анютины глазки). Одноклеточные волоски могут быть простыми (на нижней стороне многих плодовых культур), и, как правило, они мертвые. Одноклеточные волоски могут быть ветвистые (пастушья сумка). Чаще волоски бывают многоклеточными, различающимися по строению: линейными (листья картофеля), кустисто-ветвистыми (коровяк), чешуйчатыми и звездчато-чешуйчатыми (представители семейства Лоховые), массивными (пучки волосков растений семейства Губоцветные). Встречаются железистые волоски, в которых могут накапливаться эфирные вещества (губоцветные и зонтичные растения), жгучие вещества (крапива) и др. Жгучие волоски крапивы, шипы розы, ежевики, шипы на плодах зонтичных, дурмана, каштана и др. – своеобразные выросты, называемые эмергенцами, в формировании которых принимают участие помимо клеток эпидермы более глубокие слои клеток.
Эпиблема (ризодерма) – первичная однослойная покровная ткань корня. Образуется из наружных клеток апикальной меристемы корня вблизи корневого чехлика. Эпиблема покрывает молодые корневые окончания. Через нее осуществляется водно-минеральное питание растения из почвы. В эпиблеме много митохондрий. Клетки эпиблемы тонкостенны, с более вязкой цитоплазмой, лишены устьиц и кутикулы. Эпиблема недолговечна и постоянно обновляется за счет митотических делений.
Перидерма – сложный многослойный комплекс вторичной покровной ткани (пробка, пробковый камбий, или феллоген, и феллодерма) стеблей и корней многлетних двудольных растений и голосеменных, которые способны непрерывно утолщаться. К осени первого года жизни побеги одревесневают, что заметно по изменению их окраски от зеленой до буро-серой, т.е. произошла смена эпидермы на перидерму, способную выдержать неблагоприятные условия зимнего периода. В основе перидермы лежит вторичная меристема – феллоген (пробковый камбий), образующийся в клетках основной паренхимы, лежащей под эпидермой. Феллоген образует клетки в двух направлениях: наружу – клетки пробки, внутрь – живые клетки феллодермы. Пробка состоит из отмерших клеток, заполненных воздухом, они вытянуты в длину, плотно прилегают друг к другу, поры отсутствуют, клетки воздухо- и водонепроницаемы. Клетки пробки имеют коричневый или желтоватый цвет, который зависит от присутствия в клетках смолистых или дубильных веществ (пробковый дуб, бархат сахалинский). Пробка хороший изоляционный материал, не проводит тепла, электричества и звуки, используется для закупорки бутылок и др. Мощный слой пробки имеет пробковый дуб, виды бархата, пробковый вяз.
Чечевички – «вентиляционные» отверстия в пробке для обеспечения газо- и водообмена живых, более глубоко лежащих тканей растения с внешней средой. Внешне чечевички похожи на семена чечевицы, за что и получили свое название. Как правило, чечевички закладываются на смену устьицам. Формы и размеры чечевичек различны. В количественном отношении чечевичек намного меньше, чем устьиц. Чечевички представляют собой округлые тонкостенные бесхлорофилльные клетки с межклетниками, которые приподнимают кожицу и разрывают ее. Этот слой рыхлых слабо опробковевших паренхимных клеток, составляющих чечевичку, называется выполняющей тканью.
Корка – мощный покровный комплекс из отмерших наружных клеток перидермы. Она формируется на многолетних побегах и корнях древесных растений. У корки трещиноватая и неровная форма. Она предохраняет стволы деревьев от механических повреждений, низовых пожаров, низких температур, солнечных ожогов, проникновения болезнетворных бактерий и грибов. Растет корка за счет нарастания под ней новых слоев перидермы. У древесно-кустарниковых растений корка возникает (например, у сосны) на 8-10-м, а у дуба – на 25-30-м году жизни. Корка входит в состав коры деревьев. Снаружи она постоянно слущивается, сбрасывая с себя всевозможные споры грибов и лишайников.
4. Основные ткани. Основная ткань, или паренхима, занимает большую часть пространства между другими постоянными тканями стеблей, корней и других органов растений. Основные ткани состоят в основном из живых клеток, разнообразных по форме. Клетки тонкостенные, но иногда утолщенные и одревесневшие, с постенной цитоплазмой, простыми порами. Из паренхимы состоят кора стеблей и корней, сердцевина стеблей, корневищ, мякоть сочных плодов и листьев, она служит хранилищем питательных веществ в семенах. Выделяют несколько подгрупп основных тканей: ассимиляционную, запасающую, водоносную и воздухоносную.
Ассимиляционная ткань, или хлорофиллоносная паренхима, или хлоренхима, - ткань, в которой осуществляется фотосинтез. Клетки тонкостенны, содержат хлоропласты, ядро. Хлоропласты, как и цитоплазма, расположены постенно. Хлоренхима находится непосредственно под кожицей. В основном хлоренхима сосредоточена в листьях и молодых зеленых побегах растений. В листьях различают палисадную, или столбчатую, и губчатую хлоренхиму. Клетки палисадной хлоренхимы удлиненные, цилиндрической формы, с очень узкими межклетниками. Губчатая хлоренхима имеет более или менее округлые рыхло расположенные клетки с большим количеством межклетников, заполненных воздухом.
Аэренхима,или воздухоносная ткань, - паренхима со значительно развитыми межклетниками в разных органах характерна для водных, прибрежно-водных и болотных растений (камыши, ситники, кубышки, рдесты, водокрасы и др.), корни и корневища которых находятся в иле, бедном кислородом. Атмосферный воздух доходит до подводных органов через фотосинтетическую систему посредством передаточных клеток. Кроме того, воздухоносные межклетники сообщаются с атмосферой с помощью своеобразных пневматод - устьиц листьев и стеблей, пневматод воздушных корней некоторых растений (монстера, филодендрон, фикус баньян и др.), щелей, отверстий, каналов, окруженных клетками-регуляторами сообщений. Аэренхима уменьшает удельный вес растения, что, вероятно, способствует поддержанию вертикального положения водных растений, а водным растениям с плавающими на поверхности воды листьями - удержанию листьев на поверхности воды.
Водоносная ткань запасает воду в листьях и стеблях суккулентных растений (кактусы, алоэ, агавы, толстянки и др.), а также растений засоленных местообитаний (солерос, биюргун, сарсазан, солянки, гребенщик, черный саксаул и др.), как правило, в аридных областях. Листья злаков также имеют крупные водоносные клетки со слизистыми веществами, удерживающими влагу. Хорошо развитые водоносные клетки имеет мох сфагнум.
Запасающие ткани - ткани, в которых в определенный период развития растения откладывают продукты обмена - белки, углеводы, жиры и др. Клетки запасающей ткани обычно тонкостенны, паренхима живая. Запасающие ткани широко представлены в клубнях, луковицах, утолщенных корнях, сердцевине стеблей, эндосперме и зародышах семян, паренхиме проводящих тканей (фасоль, ароидные), вместилищах смол и эфирных масел в листьях лавра, камфарного дерева и др. Запасающая ткань может превращаться в хлоренхиму, например, при прорастании клубней картофеля, луковиц луковичных растений.
5. Механические ткани. Механические, или опорные, ткани- это своего рода арматура, или стереом. Термин стереом происходит от греческого «стереос» - твердый, прочный. Основная функция - обеспечение сопротивления статическим и динамическим нагрузкам. В соответствии с функциями они имеют подобающее строение. У наземных растений они наиболее развиты в осевой части побега - стебле. Клетки механической ткани могут располагаться в стебле либо по периферии, либо сплошным цилиндром, либо отдельными участками в гранях стебля. В корне, который выдерживает в основном сопротивление на разрыв, механическая ткань сосредоточена в центре. Особенность строения этих клеток - сильное утолщение клеточных стенок, которые и придают тканям прочность. Наиболее хорошо развиты механические ткани у древесных растений. По строению клеток и характеру утолщений клеточных стенок механические ткани разделяют на два типа: колленхиму и склеренхиму.
Колленхима - это простая первичная опорная ткань с живыми содержимым клеток: ядром, цитоплазмой, иногда с хлоропластами, с неравномерно утолщенными клеточными стенками. По характеру утолщений и соединения клеток между собой различают три типа колленхимы: уголковую, пластинчатую и рыхлую. Если клетки утолщены только по углам, то это уголковая колленхима, а если стенки утолщены параллельно поверхности стебля и утолщение равномерное, то это пластинчатая колленхима. Клетки уголковой и пластинчатой колленхимы расположены плотно друг к другу, не образуя межклетников. Рыхлая колленхима имеет межклетники, а утолщенные клеточные стенки направлены в сторону межклетников.
Эволюционно колленхима возникла из паренхимы. Формируется колленхима из основной меристемы и находится под эпидермой на расстоянии одного или нескольких слоев от нее. В молодых стеблях побегов она располагается в виде цилиндра по периферии, в жилках крупных листьев - по обеим их сторонам. Живые клетки колленхимы способны расти в длину, не препятствуя росту молодых растущих частей растения.
Склеренхима - наиболее распространенная механическая ткань, состоящая из клеток с одревесневшими (за исключением лубяных волокон льна) и равномерно утолщенными клеточными стенками с немногочисленными щелевидными порами. Клетки склеренхимы вытянуты в длину и имеют прозенхимную форму с заостренными концами. Оболочки склеренхимных клеток по прочности близки к стали. Содержание лигнина в этих клетках повышает прочность склеренхимы. Склеренхима есть почти во всех вегетативных органах высших наземных растений. У водных ее либо совсем нет, либо она слабо представлена в погруженных органах водных растений.
Различают первичную и вторичную склеренхиму. Первичная склеренхима происходит из клеток основной меристемы - прокамбия или перицикла, вторичная - из клеток камбия. Различают два типа склеренхимы: склеренхимные волокна, состоящие из мертвых толстостенных клеток с заостренными концами, с одревесневшей оболочкой и немногочисленными порами, как у лубяных и древесинных волокон, или волокон либроформа, и склереиды - структурные элементы механической ткани, располагающиеся в одиночку или группами между живыми клетками разных частей растения: кожуры семян, плодов, листьев, стеблей. Основная функция склереид - противостоять сдавливанию. Форма и размеры склереид разнообразны.
6. Проводящие ткани. Проводящие ткани транспортируют питательные вещества в двух направлениях. Восходящий (транспирационный) ток жидкости (водные растворы и соли) идет по сосудам и трахеидам ксилемы от корней вверх по стеблю к листьям и другим органам растения. Нисходящий ток (ассимиляционный) органических веществ осуществляется от листьев по стеблю к подземным органам растения по специальным ситовидным трубкам флоэмы. Проводящая ткань растения чем-то напоминает кровеносную систему человека, так как имеет осевую и радиальную сильно разветвленную сеть; питательные вещества попадают в каждую клеточку живого растения. В каждом органе растения ксилема и флоэма располагаются рядом и представлены в виде тяжей - проводящих пучков.
Существуют первичные и вторичные проводящие ткани. Первичные дифференцируются из прокамбия и закладываются в молодых органах растения, вторичные проводящие ткани более мощные, формируются из камбия.
Ксилема (древесина) представлена трахеидами и трахеями, или сосудами.
Трахеиды - вытянутые замкнутые клетки с косо срезанными зазубренными концами, в зрелом состоянии представлены мертвыми прозенхимными клетками. Длина клеток в среднем 1-4 мм. Сообщение с соседними трахеидами происходит через простые или окаймленные поры. Стенки неравномерно утолщены, по характеру утолщения стенок различают трахеиды кольчатые, спиральные, лестничные, сетчатые и пористые. У пористых трахеид всегда окаймленные поры. Спорофиты всех высших растений имеют трахеиды, а у большинства хвощевидных, плауновидных, папоротниковидных и голосеменных они служат единственными проводящими элементами ксилемы. Трахеиды выполняют две основные функции: проведение воды и механическое укрепление органа.
Трахеи,или сосуды, - главнейшие водопроводящие элементы ксилемы покрытосеменных растений. Трахеи представляют собой полые трубки, состоящие из отдельных члеников; в перегородках между члениками находятся отверстия - перфорации, благодаря которым осуществляется ток жидкости. Трахеи, как и трахеиды, - это замкнутая система: концы каждой трахеи имеют скошенные поперечные стенки с окаймленными порами. Членики трахей крупнее, чем трахеиды: в поперечнике составляют у разных видов растений от 0,1-0,15 до 0,3 - 0,7 мм. Длина трахей от нескольких метров до нескольких десятков метров (у лиан). Трахеи состоят из мертвых клеток, хотя на начальных стадиях формирования они живые. Считают, что трахеи в процессе эволюции возникли из трахеид.
Сосуды и трахеиды помимо первичной оболочки в большинстве имеют вторичные утолщения в виде колец, спиралей, лестниц и т.д. Вторичные утолщения образуются на внутренней стенке сосудов. Так, в кольчатом сосуде внутренние утолщения стенок в виде колец, находящихся на расстоянии друг от друга. Кольца расположены поперек сосуда и чуть наклонно. В спиральном сосуде вторичная оболочка наслаивается изнутри клетки в виде спирали; в сетчатом сосуде неутолщенные места оболочки выглядят в виде щелей, напоминающих ячеи сетки; в лестничном сосуде утолщенные места чередуются с неутолщенными, образуя подобие лестницы.
Трахеиды и сосуды - трахеальные элементы - распределяются в ксилеме различным образом: на поперечном срезе сплошными кольцами, образуя кольцесосудистую древесину, или рассеянно более или менее равномерно по всей ксилеме, образуя рассеянно-сосудистую древесину. Вторичная оболочка, как правило, пропитывается лигнином, придавая растению дополнительную прочность, но в то же время ограничивая его рост в длину.
Помимо сосудов и трахеид ксилема включает лучевые элементы, состоящие из клеток, образующих сердцевинные лучи. Сердцевинные лучи состоят из тонкостенных живых паренхимных клеток, по которым питательные вещества оттекают в горизонтальном направлении. В ксилеме присутствуют также живые клетки древесинной паренхимы, которые функционируют в качестве ближнего транспорта, и служат местом хранения запасных веществ. Все элементы ксилемы происходят из камбия.
Флоэма - проводящая ткань, по которой транспортируется глюкоза и другие органические вещества - продукты фотосинтеза от листьев к местам их использования и отложения (к конусам нарастания, клубням, луковицам, корневищам, корням, плодам, семенам и др.). Флоэма также бывает первичная и вторичная. Первичная флоэма формируется из прокамбия, вторичная (луб) - из камбия. В первичной флоэме отсутствуют сердцевинные лучи и менее мощная система ситовидных элементов, нежели у трахеид. В процессе формирования ситовидной трубки в протопласте клеток - члеников ситовидной трубки появляются слизевые тельца, принимающие участие в образовании слизевого тяжа около ситовидных пластинок. На этом формирование членика ситовидной трубки заканчивается. Функционируют ситовидные трубки у большинства травянистых растений один вегетационный период и до 3-4 лет у древесно-кустарниковых растений. Ситовидные трубки состоят из ряда удлиненных клеток, сообщающихся друг с другом посредством продырявленных перегородок - ситечек. Оболочки функционирующих ситовидных трубок не одревесневают и остаются живыми. Старые клетки закупориваются так называемым мозолистым телом, а потом отмирают и под давлением на них более молодых функционирующих клеток сплющиваются.
К флоэме относится лубяная паренхима, состоящая из тонкостенных клеток, в которых откладываются запасные питательные вещества. По сердцевинным лучам вторичной флоэмы осуществляется также ближняя транспортировка органических питательных веществ - продуктов фотосинтеза.
Проводящие пучки - тяжи, образуемые, как правило, ксилемой и флоэмой. Если к проводящим пучкам примыкают тяжи механической ткани (чаще склеренхимы), то такие пучки называют сосудисто-волокнистыми. В проводящие пучки могут быть включены и другие ткани - живая паренхима, млечники и др. Проводящие пучки могут быть полными, когда присутствуют и ксилема и флоэма, и неполными, состоящими только из ксилемы (ксилем-ный, или древесинный, проводящий пучок) или флоэмы (флоэмный, или лубяной, проводящий пучок).
Проводящие пучки первоначально образовались из прокамбия. Выделяют несколько типов проводящих пучков. Часть прокамбия может сохраниться и затем превратиться в камбий, тогда пучок способен к вторичному утолщению. Это открытые пучки. Такие проводящие пучки преобладают у большинства двудольных и голосеменных растений. Растения, имеющие открытые пучки, способны разрастаться в толщину за счет деятельности камбия, причем древесинные участки примерно в три раза крупнее лубяных участков. Если при дифференци-ровке проводящего пучка из прокамбиального тяжа вся образовательная ткань полностью расходуется на формирование постоянных тканей, то пучок называется закрытым.
Закрытые проводящие пучки встречаются в стеблях однодольных растений. Древесина и луб в пучках могут иметь различное взаимное расположение. В связи с этим выделяют несколько типов проводящих пучков: коллатеральные, биколлатеральные, концентрические и радиальные. Коллатеральные, или бокобочные, - пучки, в которых ксилема и флоэма примыкают друг к другу. Биколлатеральные, или двубокобочные, - пучки, в которых к ксилеме примыкают бок о бок два тяжа флоэмы. В концентрических пучках ткань ксилемы полностью окружает ткань флоэмы или наоборот. В первом случае такой пучок называют центрофлоэмным. Центрофлоэмные пучки имеются у стеблей и корневищ некоторых двудольных и однодольных растений (бегония, щавель, ирис, многие осоковые и лилейные). Ими обладают папоротники. Существуют и промежуточные проводящие пучки между закрытыми коллатеральными и центрофлоэмными. В корнях встречаются радиальные пучки, в которых центральную часть и лучи по радиусам оставляет древесина, причем каждый луч древесины состоит из центральных более крупных сосудов, постепенно уменьшаясь по радиусам. Число лучей у разных растений неодинаково. Между древесинными лучами располагаются лубяные участки. Проводящие пучки тянутся вдоль всего растения в виде тяжей, которые начинаются в корнях и проходят вдоль всего растения по стеблю к листьям и другим органам. В листьях они называются жилками. Главная функция их - проведение нисходящего и восходящего токов воды и питательных веществ.
7. Выделительные ткани. Выделительные, или секреторные, ткани представляют собой специальные структурные образования, способные выделять из растения или изолировать в его тканях продукты метаболизма и капельно-жидкую среду. Продукты метаболизма называют секретами. Если они выделяются наружу, то это ткани наружной секреции, если остаются внутри растения, то - внутренней секреции. Как правило, это живые паренхимные тонкостенные клетки, однако по мере накопления в них секрета они лишаются протопласта и их клетки опробковевают. Образование жидких секретов связано с деятельностью внутриклеточных мембран и комплекса Гольджи, а их происхождение - с ассимиляционными, запасающими и покровными тканями. Основная функция жидких секретов заключается в защите растения от поедания животными, повреждения насекомыми или болезнетворными микроорганизмами. Ткани внутренней секреции представлены в виде клеток-идиобластов, смоляных ходов, млечников, эфиромасличных каналов, вместилищ выделений, железистых головчатых волосков, железок. В клетках-идиобластах часто содержатся кристаллы щавелевокислого кальция (представители семейства Лилейные, Крапивные и др.), слизи (представители семейства Мальвовые и др.), терпеноиды (представители семейств Магнолиевые, Перечные и др.) и т.п.
15. Корни, их виды. Типы корневых систем. Функции корня.
Функции корня . Корень - основной орган высшего растения. Функции корней следующие:
Всасывают из почвы воду и растворенные в ней минеральные соли, транспортирует их вверх по стеблю, листьям и репродуктивным органам. Функцию всасывания выполняют корневые волоски (или микоризы), расположенные в зоне всасывания.
Благодаря большой прочности закрепляют растение в почве.
При взаимодействии воды, ионов минеральных солей и продуктов фотосинтеза синтезируют продукты первичного и вторичного метаболизма.
Под действием корневого давления и транспирации ионы водных растворов минеральных веществ и органические вещества по сосудам ксилемы корня передвигаются по восходящему току в стебель и листья.
В корнях откладываются в запас питательные вещества (крахмал, инулин и др.).
В корнях осуществляется биосинтез вторичных метаболитов (алкалоиды, гормоны и другие БАВ).
Синтезированные в меристематических зонах корней ростовые вещества (гиббереллины и др.) необходимы для роста и развития надземных частей растения.
За счет корней осуществляется симбиоз с почвенными микроорганизмами -бактериями и грибами.
С помощью корней происходит вегетативное размножение многих растений.
Некоторые корни выполняют функцию дыхательного органа (монстера, филодендрон и др.).
Корни ряда растений выполняют функцию "ходульных" корней (фикус баньян, панданус и др.).
Корень способен к метаморфозам (утолщения главного корня образуют "корнеплоды" у моркови, петрушки и др.; утолщения боковых или придаточных корней образуют корневые клубни у георгин, земляных орешков, чистяка и др., укорачивание корней у луковичных растений).
Корень - осевой орган, обычно цилиндрической формы, с радиальной симметрией, обладающий геотропизмом. Растет до тех пор, пока сохраняется верхушечная меристема, покрытая корневым чехликом. На корне в отличие от побега никогда не образуются листья, зато, как и побег, корень ветвится, образуя корневую систему .
Корневая система - это совокупность корней одного растения. Характер корневой системы зависит от соотношения роста главного, боковых и придаточных корней.
Типы корней и корневых систем . В зародыше семени все органы растения находятся в зачаточном состоянии. Главный, или первый, корень развивается из зародышевого корешка . Главный корень расположен в центре всей корневой системы, стебель служит продолжением корня, и вместе они составляют ось первого порядка. Участок на границе между главным корнем и стеблем называется корневой шейкой . Этот переход от стебля к корню заметен по разной толщине стебля и корня: стебель более толстый, чем корень. Участок стебля от корневой шейки до первых зародышевых листьев - семядолей называют подсемядольным коленом или гипокотилем (рис. 46). От главного корня в стороны отходят боковые корни очередных порядков. Такая корневая система называется стержневой (рис. 47, А, Б) , у многих двудольных растений она способна к ветвлению. Ветвистая корневая система - разновидность стержневой корневой системы. Боковое ветвление корня характеризуется тем, что новые корни закладываются на некотором расстоянии от верхушки и образуются эндогенно - во внутренних тканях материнского корня предшествующего порядка за счет активности перицикла. Чем больше отходит боковых корней от главного корня, тем больше площадь питания растения, поэтому существуют специальные агротехнические приемы, усиливающие способность главного корня образовывать боковые, например прищипывание или пикирование главного корня на l/3 его длины. После пикирования некоторое время главный корень перестает расти в длину, в то время как боковые корни интенсивно растут.
У двудольных растений главный корень, как правило, сохраняется всю жизнь, у однодольных зародышевый корешок быстро отмирает, главный корень не развивается, а от основания побега образуются придаточные корни (рис. 47, Г) , которые также имеют ответвления первого, второго и т.д. порядков. Такая корневая система называется мочковатой (рис. 47, В, Г) . Придаточные корни, как и боковые, закладываются эндогенно. Они могут образовываться на стеблях и листьях. Способность растений развивать придаточные корни широко используется в растениеводстве при вегетативном размножении растений (размножение стеблевыми и листовыми черенками). Надземными стеблевыми черенками размножают иву, тополь, клен, смородину черную и др.; листовыми черенками - фиалку узамбарскую, или сенполию, некоторые виды бегоний. Подземными черенками видоизмененных побегов (корневищ) размножают многие лекарственные растения, например, ландыш майский, купену лекарственную и др. Некоторые растения образуют много придаточных корней при окучивании нижней части стебля (картофель, капуста, кукуруза и др.), создавая тем самым дополнительное питание.
У высших споровых растений (плауны, хвощи, папоротники) главный корень вообще не возникает, у них формируются только придаточные корни, отходящие от корневища. У многих двудольных травянистых корневищных растений главный корень часто отмирает и преобладает система придаточных корней, отходящих от корневищ (сныть, крапива, лютик ползучий и др.).
По глубине проникновения в почву первое место принадлежит стержневой корневой системе: рекордная глубина проникновения корней, по некоторым сведениям, достигает 120 м! Однако мочковатая корневая система, обладая в основном поверхностным расположением корней, способствует созданию дернового покрова и предупреждает эрозию почвы.
Общая длина корней в корневой системе различна, некоторые корни достигают нескольких десятков и даже сотен километров. Например, у пшеницы длина всех корневых волосков достигает 20 км, а у озимой ржи общая длина корней первого, второго и третьего порядков свыше 180 км, а с присоединением корней четвертого порядка - 623 км. Несмотря на то что корень растет всю жизнь, рост его ограничен влиянием корней других растений.
Степень развития корневых систем на разных почвах в разных природных зонах неодинакова. Так, в песчаных пустынях, где глубоко залегают грунтовые воды, корни некоторых растений уходят на глубину 40 м и более (злак селин, просопис сережкоцветный из семейства Бобовые и др.). Растения-эфемеры полупустынь имеют поверхностную корневую систему, которая приспособлена к быстрому впитыванию ранневесенней влаги, вполне достаточной для быстрого прохождения всех фаз вегетации растений. На глинистых, плохо аэрируемых подзолах таежной лесной зоны корневая система растений на 90 % сосредоточена в поверхностном слое почвы (10- 15 см), растения имеют "питающие корни" (ель европейская). Например, у саксаула корни в разное время года используют влагу разных горизонтов.
Очень важный фактор в распределении корневой системы - влажность. Направление корней идет в сторону большей влажности, однако в воде и в переувлажненной почве корни ветвятся намного слабее.
Химический состав клетки. Неорганические вещества клетки
Основные свойства и уровни организации живой природы
Уровни организации живых систем отражают соподчиненность, иерархичность структурной организации жизни:
Молекулярно-генетический - отдельные биополимеры (ДНК, РНК, белки);
Клеточный - элементарная самовоспроизводящаяся единица жизни (прокариоты, одноклеточные эукариоты), ткани, органы;
Организменный - самостоятельное существование отдельной особи;
Популяционно-видовой - элементарная эволюционирующая единица - популяция;
Биогеоценотический - экосистемы, состоящие из разных популяций и среды их обитания;
Биосферный - все живое население Земли, обеспечивающее круговорот веществ в природе.
Природа - это весь существующий материальный мир во всем многообразии его форм. Единство природы проявляется в объективности ее существования, общности элементного состава, подчиненности одним и тем же физическим законам, в системности организации. Различные природные системы, как живые, так и неживые, взаимосвязаны и взаимодействуют между собой. Примером системного взаимодействия является биосфера.
Биология - это комплекс наук, изучающих закономерности развития и жизнедеятельности живых систем, причины их многообразия и приспособленности к окружающей среде, взаимосвязь с другими живыми системами и объектами неживой природы.
Объектом исследования биологии является живая природа.
Предметом исследования биологии являются:
Общие и частные закономерности организации, развития, обмена веществ, передачи наследственной информации;
Разнообразие форм жизни и самих организмов, а также их связи с окружающей средой.
Все многообразие жизни на Земле объясняется эволюционным процессом и действием окружающей среды на организмы.
Сущность жизни определяется М.В. Волькенштейном как существование на Земле «живых тел, представляющих собой открытые саморегулирующиеся и самовоспроизводящиеся системы, построенные из биополимеров - белков и нуклеиновых кислот».
Основные свойства живых систем:
Обмен веществ;
Саморегуляция;
Раздражимость;
Изменчивость;
Наследственность;
Размножение;
Химический состав клетки. Неорганические вещества клетки
Цитология - наука, изучающая строение и функции клеток. Клетка является элементарной структурной и функциональной единицей живых организмов. Клеткам одноклеточных организмов присущи все свойства и функции живых систем. Клетки многоклеточных организмов дифференцированы по строению и функциям.
Атомный состав: в состав клетки входит около 70 элементов Периодической системы элементов Менделеева, причем 24 из них присутствуют во всех типах клеток.
Макроэлементы - Н, О, N, С, микроэлементы - Mg, Na, Са, Fe, К, Р, CI, S, ультрамикроэлементы - Zn, Сu, I, F, Мn, Со, Si и др.
Молекулярный состав: в состав клетки входят молекулы неорганических и органических соединений.
Неорганические вещества клетки
Вода. Молекула воды имеет нелинейную пространственную структуру и обладает полярностью. Между отдельными молекулами образуются водородные связи, определяющие физические и химические свойства воды.
Рис. 1. Молекула воды Рис. 2. Водородные связи между молекулами воды
Физические свойства воды:
Вода может находиться в трех состояниях - жидком, твердом и газообразном;
Вода - растворитель. Полярные молекулы воды растворяют полярные молекулы других веществ. Вещества, растворимые в воде, называют гидрофильными. Вещества, не растворимые в воде, - гидрофобными;
Высокая удельная теплоемкость. Для разрыва водородных связей, удерживающих молекулы воды, требуется поглотить большое количество энергии. Это свойство воды обеспечивает поддержание теплового баланса в организме;
Высокая теплота парообразования. Для испарения воды необходима достаточно большая энергия. Температура кипения воды выше, чем у многих других веществ. Это свойство воды предохраняет организм от перегрева;
Молекулы воды находятся в постоянном движении, они сталкиваются друг с другом в жидкой фазе, что немаловажно для процессов обмена веществ;
Сцепление и поверхностное натяжение. Водородные связи обусловливают вязкость воды и сцепление ее молекул с молекулами других веществ (когезия). Благодаря силам сцепления молекул на поверхности воды создается пленка, которую характеризует поверхностное натяжение;
Плотность. При охлаждении движение молекул воды замедляется. Количество водородных связей между молекулами становится максимальным. Наибольшую плотность вода имеет при 4°С. Замерзая, вода расширяется (необходимо место для образования водородных связей), и ее плотность уменьшается, поэтому лед плавает на поверхности воды, что защищает водоем от промерзания;
Способность к образованию коллоидных структур. Молекулы воды образуют вокруг нерастворимых молекул некоторых веществ оболочку, препятствующую образованию крупных частиц. Такое состояние этих молекул называется дисперсным (рассеянным). Мельчайшие частицы веществ, окруженные молекулами воды, образуют коллоидные растворы (цитоплазма, межклеточные жидкости).
Биологические функции воды:
Транспортная - вода обеспечивает передвижение веществ в клетке и организме, поглощение веществ и выведение продуктов метаболизма. В природе вода переносит продукты жизнедеятельности в почвы и к водоемам;
Метаболическая - вода является средой для всех биохимических реакций и донором электронов при фотосинтезе, она необходима для гидролиза макромолекул до их мономеров;
Участвует в образовании:
1) смазывающих жидкостей, которые уменьшают трение (синовиальная - в суставах позвоночных животных, плевральная, в плевральной полости, перикардиальная - в околосердечной сумке);
2) слизей, которые облегчают передвижение веществ по кишечнику, создают влажную среду на слизистых оболочках дыхательных путей;
3) секретов (слюна, слезы, желчь, сперма и т.д.) и соков в организме.
Неорганические ионы. Неорганические ионы клетки представлены: катионами К+, Na+, Са2+, Mg2+, NH3 и анионами Сl-, NOi2-, H2PO4-, HCO3-, HPO42-.
Разность между количеством катионов и анионов на поверхности и внутри клетки обеспечивает возникновение потенциала действия, что лежит в основе нервного и мышечного возбуждения.
Анионы фосфорной кислоты создают фосфатную буферную систему, поддерживающую рН внутриклеточной среды организма на уровне 6-9.
Угольная кислота и ее анионы создают бикарбонатную буферную систему и поддерживают рН внеклеточной среды (плазмы крови) на уровне 4-7.
Соединения азота служат источником минерального питания, синтеза белков, нуклеиновых кислот. Атомы фосфора входят в состав нуклеиновых кислот, фосфолипидов, а также костей позвоночных, хитинового покрова членистоногих. Ионы кальция входят в состав вещества костей, они также необходимы для осуществления мышечного сокращения, свертывания крови.


